Seebio(R) 煙酰胺單核苷酸應(yīng)用
Seebio®煙酰胺單核苷酸(nicotinamide mononucleotide,NMN)是一種自然存在的生物活性核苷酸,NMN有2種不規(guī)則存在形式,α和β;β異構(gòu)體是NMN的活性形式,分子量為334.2g/mol。
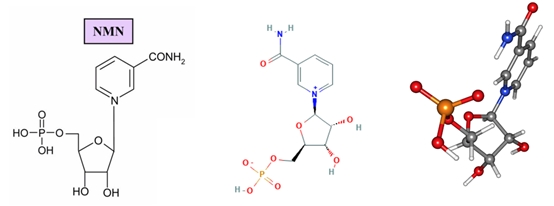
煙酰胺單核苷酸化學(xué)結(jié)構(gòu)式和球棍模型
在細(xì)胞內(nèi),尼克酰胺單核苷酸(NMN)是煙酰胺腺嘌呤二核苷酸(NAD+)的重要前體。NAD+是一種在細(xì)胞內(nèi)參與多種生物化學(xué)反應(yīng)的輔酶,對(duì)于維持細(xì)胞正常功能和代謝至關(guān)重要。因此,NMN在細(xì)胞內(nèi)具有重要的生物學(xué)意義。
|
產(chǎn)品名稱(chēng)
|
CAS
|
純度
|
包裝
|
|
β-煙酰胺單核苷酸 (NMN)
|
1094-61-7
|
99%
|
1kg 5kg 10kg 25kg
|
詳詢西寶生物產(chǎn)品,請(qǐng)咨詢:400-021-8158 / 021-50272975。
NMN對(duì)NAD+的促進(jìn)作用
主要體現(xiàn)在其作為NAD+的前體的功能上。在合成途徑中,煙酰胺核糖或煙酰胺通過(guò)NRK(煙酰胺核苷激酶)或NAMPT、NMNAT合成煙酰胺單核苷酸(NMN),隨后,NMN通過(guò)NMNAT1-3酶合成NAD+。

盡管不能在血清中檢測(cè)到完整結(jié)構(gòu)的NMN,但攝入NMN仍能夠在很短的時(shí)間內(nèi)(15分鐘)顯著提高雌性和雄性小鼠的NAD+水平。
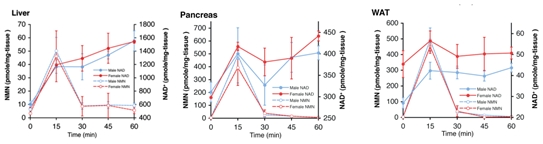
肝臟、胰腺、白色脂肪組織NMN、NAD+水平
NMN功效
NMN的主要作用是通過(guò)轉(zhuǎn)化為NAD+發(fā)揮其生物學(xué)功能。NAD+,全名煙酰胺腺嘌呤雙核苷酸,又被稱(chēng)為輔酶I,廣泛分布在人體的所有細(xì)胞內(nèi),參與上千種生物催化反應(yīng),是人體內(nèi)不可或缺的輔酶。
在衰老過(guò)程中,NAD+水平的下降被認(rèn)為是導(dǎo)致各種疾病和殘疾的主要原因,包括抗衰老、認(rèn)知和運(yùn)動(dòng)功能障礙、免疫缺陷,以及由自身免疫炎癥反應(yīng)失調(diào)引起的關(guān)節(jié)炎、代謝障礙和心血管疾病。
通過(guò)補(bǔ)充N(xiāo)MN來(lái)提高體內(nèi)NAD+含量,有助于延緩、改善和預(yù)防與衰老相關(guān)的多種表型,包括年齡誘導(dǎo)的代謝紊亂和老年疾病。

NAD+與衰老
人體持續(xù)進(jìn)行能量合成與代謝,以保持生命活動(dòng)的平衡與健康。在這一過(guò)程中,煙酰胺腺嘌呤二核苷酸(NAD+)作為氧化還原反應(yīng)的輔酶,扮演著關(guān)鍵角色。NAD+直接或間接地調(diào)控多個(gè)細(xì)胞功能,包括線粒體內(nèi)的能量合成、DNA損傷修復(fù)、染色質(zhì)重塑,以及細(xì)胞衰老和免疫細(xì)胞功能。這些細(xì)胞的基本生理過(guò)程和功能對(duì)于維持組織、代謝動(dòng)態(tài)平衡及健康衰老至關(guān)重要。
衰老是人體的自然過(guò)程。隨著年齡增長(zhǎng),人體內(nèi)NAD+水平逐漸減少,導(dǎo)致多個(gè)器官內(nèi)的線粒體能量合成下降,包括大腦、脂肪組織、皮膚、肝臟、骨骼肌和胰腺。與NAD+水平的降低相關(guān)聯(lián)的是一系列與年齡有關(guān)的疾病,如認(rèn)知功能減退、癌癥、代謝性疾病、肌少癥和衰弱。因此,通過(guò)調(diào)節(jié)NAD+代謝通路,增加NAD+水平可能成為改善衰老相關(guān)疾病、延長(zhǎng)人類(lèi)壽命和健康壽命的潛在途徑。
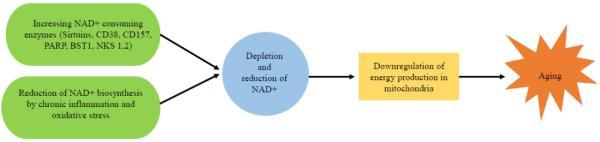
衰老過(guò)程中體內(nèi)NAD+減少的原因
NMN與抗衰老
作為NAD+的前體物質(zhì),NMN理論上通過(guò)增加體內(nèi)NAD+水平,減緩隨年齡增長(zhǎng)而導(dǎo)致的NAD+枯竭過(guò)程,從而成為一種抗衰老保健品。
科學(xué)界通過(guò)細(xì)胞試驗(yàn)、動(dòng)物模型和臨床試驗(yàn)對(duì)NMN的抗衰老活性進(jìn)行了廣泛研究。研究發(fā)現(xiàn),NMN在嚙齒動(dòng)物模型和人體水平上均表現(xiàn)出有益效果,如保護(hù)內(nèi)皮細(xì)胞、改善血供、改善代謝功能障礙以及保護(hù)神經(jīng)系統(tǒng)。一項(xiàng)2021年的隨機(jī)雙盲臨床試驗(yàn)顯示,NMN治療對(duì)絕經(jīng)后超重或肥胖的糖尿病前期女性產(chǎn)生了積極效果,增強(qiáng)了肌肉對(duì)胰島素的敏感性。類(lèi)似的研究在2023年也表明,NMN治療能夠顯著提高血清煙酰胺水平,減緩動(dòng)脈僵硬度。此外,NMN的長(zhǎng)期使用和潛在的副作用需要更多深入的研究和驗(yàn)證。
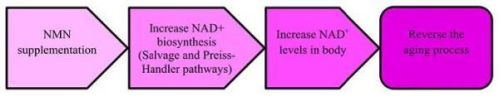
NMN抗衰老的機(jī)制
NAD+與神經(jīng)系統(tǒng)
Sirtuins是依賴NAD+的脫酰基酶,傳統(tǒng)上與熱量限制和衰老有關(guān)。在神經(jīng)發(fā)育中,SIRT1通過(guò)Akt-GSK3通路促進(jìn)軸索生長(zhǎng)、神經(jīng)突生長(zhǎng)和樹(shù)枝狀分支。在突觸的發(fā)育和調(diào)節(jié)中,sirtuins蛋白在生理和損傷后都發(fā)揮著重要的調(diào)節(jié)作用。特別是在海馬體中,SIRT1以抑制型復(fù)合體形式存在,參與調(diào)控microRNA-134的轉(zhuǎn)錄因子YY1,這對(duì)于突觸的形成和長(zhǎng)期的增強(qiáng)至關(guān)重要。
在神經(jīng)疾病的發(fā)展中,SIRT1發(fā)揮著保護(hù)作用,尤其在阿爾茨海默氏病、帕金森氏病和運(yùn)動(dòng)神經(jīng)元病等神經(jīng)退行性疾病中。這些保護(hù)作用可能與SIRT1在代謝、抗應(yīng)激和基因組穩(wěn)定性方面的功能有關(guān)。因此,激活SIRT1的藥物可能為治療這些神經(jīng)疾病提供一種有希望的方法。
NAD+與癌癥
研究表明增加NAD+水平可能對(duì)癌癥治療具有潛在效果。NMNAT3的過(guò)表達(dá)可以提高線粒體NAD+水平,從而抑制膠質(zhì)母細(xì)胞瘤細(xì)胞的生長(zhǎng)。補(bǔ)充煙酰胺(NA)或煙酰胺單核苷酸(NAM)可以抑制SCID小鼠的腫瘤生長(zhǎng)和多器官腫瘤轉(zhuǎn)移。其原理包括過(guò)量的NAD+促進(jìn)線粒體呼吸,降低糖酵解,抵消癌細(xì)胞傾向的Warburg代謝。增加NAD+還能活化SIRT1和SIRT6,兩者通過(guò)下調(diào)β-catenin信號(hào)和抑制糖酵解來(lái)抑制腫瘤。然而存在一些矛盾和擔(dān)憂,如NAD+促進(jìn)DNA修復(fù)和血管生成,可能有助于癌細(xì)胞的生長(zhǎng)。此外,降低腫瘤NAD+水平可能增加癌細(xì)胞對(duì)化療藥物的敏感性。因此,在標(biāo)準(zhǔn)癌癥模型中進(jìn)一步測(cè)試NAD+補(bǔ)充劑的效果是非常重要的。
NAD+與晝夜節(jié)律
NAD+是一種關(guān)鍵的生物分子,其依賴的脫乙酰酶SIRT1通過(guò)連接調(diào)節(jié)NAD+補(bǔ)救途徑的酶反饋回路和晝夜節(jié)律轉(zhuǎn)錄-翻譯反饋回路,成為晝夜節(jié)律與代謝之間的橋梁。NAD+通過(guò)SIRT1實(shí)現(xiàn)對(duì)生物鐘的調(diào)節(jié),其中SIRT1將BMAL1和PER2去乙酰化,抑制CLOCK-BMAL1介導(dǎo)的clock genes的轉(zhuǎn)錄。此外,NAD+通過(guò)影響SIRT1的去乙酰化活性,反過(guò)來(lái)影響包括NAMPT在內(nèi)的一系列生物鐘相關(guān)蛋白的表達(dá)。
生物鐘調(diào)節(jié)與多種疾病相關(guān),包括但不限于睡眠障礙、糖尿病和腫瘤。紊亂的生物鐘可能是多種病理過(guò)程的觸發(fā)因素,可能源自遺傳或環(huán)境因素。總體而言,維持正常的生物鐘功能對(duì)于保持健康至關(guān)重要。
NAD+與肝功能
NAD+信號(hào)通路中的酶已被證明可以保護(hù)肝臟免受脂肪堆積、纖維化和胰島素抵抗的影響,這些因素與脂肪肝病發(fā)生有關(guān)。NAMPT在高脂飲食誘導(dǎo)的脂肪肝發(fā)生中發(fā)揮關(guān)鍵調(diào)節(jié)作用,其抑制將使肝脂肪變性更加嚴(yán)重,而NAMPT的過(guò)表達(dá)則改善肝脂質(zhì)積累。SIRT1及其下游靶點(diǎn)(PGC-1a、PSK9和SREBP1)維持線粒體功能、膽固醇轉(zhuǎn)運(yùn)和脂肪酸穩(wěn)態(tài)。SIRT2通過(guò)去乙酰化磷酸烯醇丙酮酸羧激酶來(lái)控制糖異生,SIRT3調(diào)控OXPHOS、脂肪酸氧化、酮生成和抗氧化應(yīng)激,而SIRT6控制糖異生。由于這些通路在肝臟中的重要性,維持NAD+水平對(duì)于維持器官功能至關(guān)重要。在正常情況下,由于肥胖和衰老,NAMPT水平下降,CD38水平升高,導(dǎo)致到中年時(shí),穩(wěn)態(tài)NAD+水平下降2倍。將NAD+水平提高到年輕水平在預(yù)防和治療肥胖、酒精性脂肪性肝炎和NASH方面表現(xiàn)出顯著效果,同時(shí)還改善葡萄糖穩(wěn)態(tài)和線粒體功能,增強(qiáng)肝臟再生能力,保護(hù)肝臟免受肝毒性損害。
NAD+與腎功能
老年腎臟中NAD+水平的降低和sirtuin活性的下降在很大程度上是腎功能和順應(yīng)性隨年齡下降的原因。通過(guò)NAD+補(bǔ)充可以激活SIRT1和SIRT3,從而保護(hù)高糖誘導(dǎo)的腎系膜細(xì)胞肥大。同時(shí),使用NMN治療小鼠以SIRT1依賴的方式保護(hù)順鉑誘導(dǎo)的急性腎損傷(AKI)。5-氨基咪唑-4-羧胺核苷可刺激AMPK活性,增加NAD+水平,并以SIRT3依賴的方式保護(hù)順鉑誘導(dǎo)的AKI。小鼠補(bǔ)充N(xiāo)AM可以刺激腎臟保護(hù)前列腺素PGE2的分泌,提升缺血后腎功能;同時(shí),NAM也通過(guò)刺激NAD+合成抑制順鉑誘導(dǎo)的AKI。
NAD+與骨骼肌
與年輕的野生型小鼠相比,老年小鼠的肌肉表現(xiàn)出萎縮、炎癥標(biāo)志物增加以及胰島素信號(hào)和胰島素刺激葡萄糖攝取能力下降。有關(guān)骨骼肌的研究表明:用NAD+前體治療老年小鼠可以顯著改善肌肉功能。使用NMN進(jìn)行治療可以通過(guò)增加線粒體功能、提高ATP生成、減少炎癥,并將糖酵解II型肌肉轉(zhuǎn)變?yōu)檠趸w維型肌肉,逆轉(zhuǎn)與年齡相關(guān)的有害變化。這表明NAD+的補(bǔ)充可能有助于保持肌肉健康和功能,特別是在老年階段。
NAD+與心臟功能
NAD+水平對(duì)正常心臟功能和損傷后的恢復(fù)至關(guān)重要。有關(guān)心臟功能的研究表明:SIRT3在心臟功能中是至關(guān)重要的,SIRT3敲除小鼠表現(xiàn)為OXPHOS酶的高度乙酰化,導(dǎo)致ATP減少,使主動(dòng)脈對(duì)收縮高度敏感,可能與線粒體通透性過(guò)渡孔的調(diào)節(jié)因子CypD的激活有關(guān)。SIRT3-KO鼠在13個(gè)月大時(shí)出現(xiàn)纖維化和心肌肥厚,隨著年齡增長(zhǎng),病情進(jìn)一步加劇,而NMN治療可以逆轉(zhuǎn)這種下降。使用NAMPT過(guò)表達(dá)或NMN治療能顯著防止壓力過(guò)載和缺血-再灌注損傷,減少梗死面積。NAD+前體治療改善了老年MDX心肌病小鼠的心臟功能。NAD+前體提高了缺鐵誘導(dǎo)的心力衰竭小鼠模型的線粒體和心臟功能。NAD+前體通過(guò)激活SIRT3保護(hù)并恢復(fù)弗里德希氏共濟(jì)失調(diào)(FRDA)心肌病小鼠模型的心臟功能。
NAD+與血管內(nèi)皮細(xì)胞
內(nèi)皮細(xì)胞(EC)的衰老與血管疾病密切相關(guān),而NAD+前體NMN的補(bǔ)充在一些研究中顯示了改善效果:NMN治療老年小鼠可恢復(fù)頸動(dòng)脈內(nèi)皮依賴性擴(kuò)張,降低主動(dòng)脈脈搏波速和彈性動(dòng)脈剛度。NMN對(duì)老年小鼠的治療取得顯著療效,通過(guò)促進(jìn)SIRT1-依賴的毛細(xì)血管密度的增加,改善了老年小鼠的血液流動(dòng)和耐力。NMN通過(guò)改善老年小鼠的血管內(nèi)皮功能和神經(jīng)血管耦合反應(yīng),顯著提高老年小鼠的認(rèn)知。同時(shí),NMN還降低了老齡鼠腦微血管內(nèi)皮細(xì)胞的線粒體ROS,恢復(fù)了NAD+和線粒體能量。在血管內(nèi)皮中增加NAD+水平可能成為一種提高老年人活動(dòng)能力的潛在療法,并能治療因血流減少而導(dǎo)致的疾病,如缺血-再灌注損傷、傷口愈合緩慢、肝功能障礙和肌肉肌病等。
NAD+與代謝障礙
NMN對(duì)脂肪代謝和糖代謝紊亂導(dǎo)致的肥胖、Ⅱ型糖尿病、生殖抑制都具有改善作用。甚至能夠改善肥胖母親對(duì)雌性后代生殖的不良影響。 NAD+前體的補(bǔ)充可能在代謝障礙的治療中發(fā)揮重要作用。
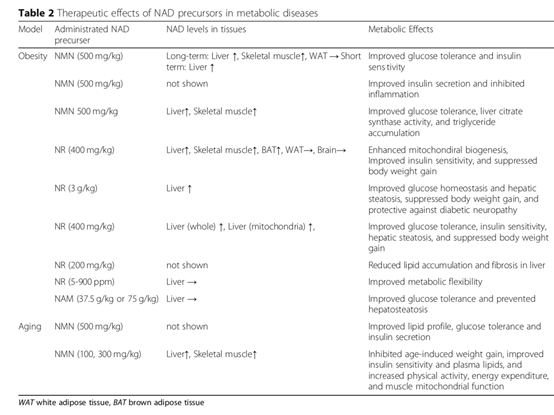
NAD+前體在動(dòng)物實(shí)驗(yàn)中對(duì)代謝障礙的治療作用
參考文獻(xiàn):
[1].Nadeeshani, H., et al., Nicotinamide mononucleotide (NMN) as an anti-aging health product - Promises and safety concerns. J Adv Res, 2022. 37: 267-278.
[2]. Liu, L., Su, X., Quinn, W. J., 3rd, Hui, S., Krukenberg, K., Frederick, D. W., . . . Rabinowitz, J. D. (2018). Quantitative Analysis of NAD Synthesis-Breakdown Fluxes. Cell Metab, 27(5), 1067-1080 e1065. doi:10.1016/j.cmet.2018.03.018
[3]. Bonkowski, M. S., & Sinclair, D. A. (2016). Slowing ageing by design: the rise of NAD(+) and sirtuin-activating compounds. Nat Rev Mol Cell Biol, 17(11), 679-690. doi:10.1038/nrm.2016.93
[4]. Clement, J., Wong, M., Poljak, A., Sachdev, P., & Braidy, N. (2019). The Plasma NAD(+) Metabolome Is Dysregulated in "Normal" Aging. Rejuvenation Res, 22(2), 121-130. doi:10.1089/rej.2018.2077
[5]. Yang, Y., & Sauve, A. A. (2016). NAD(+) metabolism: Bioenergetics, signaling and manipulation for therapy. Biochim Biophys Acta, 1864(12), 1787-1800. doi:10.1016/j.bbapap.2016.06.014
[6].H. Kim, E.L. Jacobson, M.K. Jacobson, Science 261 (1993) 1330–1333.
[7]. Pehar, M., Harlan, B. A., Killoy, K. M., & Vargas, M. R. (2018). Nicotinamide Adenine Dinucleotide Metabolism and Neurodegeneration. Antioxid Redox Signal, 28(18), 1652-1668. doi:10.1089/ars.2017.7145
[8].M.B. Schultz, D.A. Sinclair, Cell Metab. 23 (2016) 965–966.
[9]. Davila, A., Liu, L., Chellappa, K., Redpath, P., Nakamaru-Ogiso, E., Paolella, L. M., . . . Baur, J. A. (2018). Nicotinamide adenine dinucleotide is transported into mammalian mitochondria. Elife, 7. doi:10.7554/eLife.33246
[10].M. Howard, J.C. Grimaldi, J.F. Bazan, F.E. Lund, L. Santos-Argumedo, R.M. Parkhouse, T.F. Walseth, H.C. Lee, Science 262 (1993) 1056–1059.[6
[11].J. Camacho-Pereira, M.G. Tarragó, C.C.S. Chini, V. Nin, C. Escande, G.M. Warner, A.S. Puranik, R.A. Schoon, J.M. Reid, A. Galina, E.N. Chini, Cell Metabolism 23 (2016) 1127–1139.
[12]. Chini, C. C. S., Tarrago, M. G., & Chini, E. N. (2017). NAD and the aging process: Role in life, death and everything in between. Mol Cell Endocrinol, 455, 62-74. doi:10.1016/j.mce.2016.11.003
[13]. Grozio, A., Mills, K. F., Yoshino, J., Bruzzone, S., Sociali, G., Tokizane, K., . . . Imai, S. I. (2019). Slc12a8 is a nicotinamide mononucleotide transporter. Nat Metab, 1(1), 47-57. doi:10.1038/s42255-018-0009-4
[14]. Araki, T., Sasaki, Y., & Milbrandt, J. (2004). Increased nuclear NAD biosynthesis and SIRT1 activation prevent axonal degeneration. Science, 305(5686), 1010-1013. doi:10.1126/science.1098014
[15]. Ratajczak, J., Joffraud, M., Trammell, S. A., Ras, R., Canela, N., Boutant, M., . . . Canto, C. (2016). NRK1 controls nicotinamide mononucleotide and nicotinamide riboside metabolism in mammalian cells. Nat Commun, 7, 13103. doi:10.1038/ncomms13103
[16]. Pehar, M., Harlan, B. A., Killoy, K. M., & Vargas, M. R. (2018). Nicotinamide Adenine Dinucleotide Metabolism and Neurodegeneration. Antioxid Redox Signal, 28(18), 1652-1668. doi:10.1089/ars.2017.7145
[17].J. Clement, M. Wong, A. Poljak, P. Sachdev, N. Braidy, Rejuvenation Res. (2018).
二維碼: www.baichuan365.com") |
號(hào):iseebio") |
 |
| 官網(wǎng):www.baichuan365.com | 微信服務(wù)號(hào):iseebio | 微博:seebiobiotech |
 |
:seebiotech") |
堂官網(wǎng):www.canmedo.com") |
| 商城:mall.seebio.cn | 微信訂閱號(hào):seebiotech | 泉養(yǎng)堂:www.canmedo.com |
相關(guān)資訊
- Cell:體內(nèi)唾液酸化作用可將有害抗體變?yōu)橛幸婵寡卓贵w
- Cell:鑒定出彌漫大B細(xì)胞淋巴瘤的驅(qū)動(dòng)基因
- 天野脂肪酶Amano Lipase-符合“綠色化學(xué)”觀念的生物催化劑
- 乳糖/D-半乳糖[快速]檢測(cè)試劑盒
- 西寶生物 Jackson Immuno Research - 代理證書(shū)
- 科學(xué)家發(fā)現(xiàn)抗高溫基因 水稻“高溫不死”還能增產(chǎn)
- 增塑劑讓快餐不健康 我們還是去“吃土”吧!
- Nature:重磅!一些人胚胎干細(xì)胞系發(fā)生癌癥相關(guān)突變
- PNAS:解讀!科學(xué)家們有望利用一種新型的聚糖標(biāo)記策略來(lái)揭秘肝臟中的免疫激活機(jī)制!
- Culture(R) A-83-01

")
新進(jìn)產(chǎn)品
同類(lèi)文章排行
- 羥丙基殼聚糖:性能提升與多領(lǐng)域應(yīng)用
- “雜草”變身寶貝——油莎草根油 沙漠中的軟黃金
- 青刺果油:源自自然的保濕抗衰產(chǎn)品
- 蝦青素功效及應(yīng)用場(chǎng)景
- 玻色因原料: 全方位修復(fù)皮膚的抗衰劑
- 谷胱甘肽功效及應(yīng)用場(chǎng)景
- 輔酶Q10功效及應(yīng)用場(chǎng)景
- 花生四烯酸(Arachidonic acid)功能及應(yīng)用場(chǎng)景
- 黑曲霉糖(Nigerose)功效及應(yīng)用場(chǎng)景
- 曲二糖 (Kojibiose,CAS: 2140-29-6)性質(zhì)及應(yīng)用場(chǎng)景
資訊文章
您的瀏覽歷史